

GJ Mendel (1865) a étudié l'hérédité de caractères bien définis de la plante de pois, mais pour plusieurs raisons, il n'a pas été reconnu jusqu'en 1900. Trois scientifiques (de Vries, Correns et Tschermak) ont redécouvert indépendamment les résultats de Mendel sur l'hérédité des caractères. Divers cytologistes ont également observé la division cellulaire en raison des progrès de la microscopie. Cela a conduit à la découverte de structures à l'intérieur du noyau. Dans les cellules eucaryotes, les structures en forme de ver formées lors de la division cellulaire sont appelées chromosomes (corps colorés, visualisés par coloration). Un organisme qui possède deux ensembles de base complets de chromosomes est appelé diploïde. Un chromosome est constitué d'un long morceau d'ADN enroulé et continu dans lequel les gènes sont disposés dans un ordre linéaire. Chaque gène a une position définie (locus) sur un chromosome. Ces gènes sont des unités héréditaires. La théorie chromosomique de l'hérédité stipule que les facteurs mendéliens (gènes) ont un locus (position) spécifique sur les chromosomes et qu'ils transmettent des informations d'une génération à la génération suivante.
1. Développement historique de la théorie des chromosomes
Les découvertes cytologiques importantes liées à la théorie chromosomique de l'hérédité sont présentées ci-dessous.
Wilhelm Roux (1883) a postulé que les chromosomes d'une cellule sont responsables du transfert de l'hérédité.
Montgomery (1901) a été le premier à suggérer la présence de paires distinctes de chromosomes et il a également conclu que les chromosomes maternels ne s'apparient avec les chromosomes paternels que pendant la méiose.
T. Boveri (1902) a soutenu l'idée que les chromosomes contiennent des déterminants génétiques, et il était en grande partie responsable du développement de la théorie chromosomique de l'hérédité.
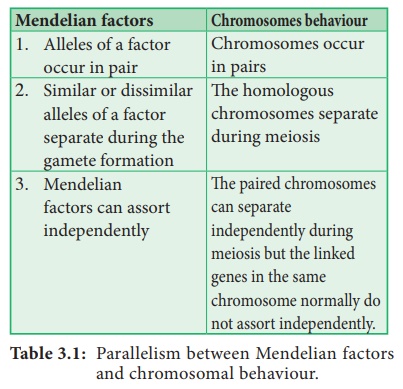
WS Sutton (1902) , un jeune étudiant américain a indépendamment reconnu un parallélisme (similarité) entre le comportement des chromosomes et les facteurs mendéliens lors de la formation des gamètes. Sutton et Boveri (1903) ont proposé indépendamment la théorie chromosomique de l'hérédité. Sutton a uni la connaissance de la ségrégation chromosomique aux principes mendéliens et l'a appelée théorie chromosomique de l'hérédité.
2. Traits saillants de la théorie chromosomique de l'hérédité
· Les cellules somatiques des organismes sont dérivées du zygote par division cellulaire répétée (mitose). Ceux-ci sont constitués de deux ensembles identiques de chromosomes. Un ensemble est reçu du parent femelle (maternel) et l'autre du parent mâle (paternel). Ces deux chromosomes constituent la paire homologue.
· Les chromosomes conservent leur unicité structurelle et leur individualité tout au long du cycle de vie d'un organisme.
· Chaque chromosome porte des déterminants spécifiques ou des facteurs mendéliens qui sont maintenant appelés gènes.
· Le comportement des chromosomes lors de la formation des gamètes (méiose) apporte la preuve que des gènes ou des facteurs sont localisés sur les chromosomes.
3. Appui à la théorie chromosomique de l'hérédité
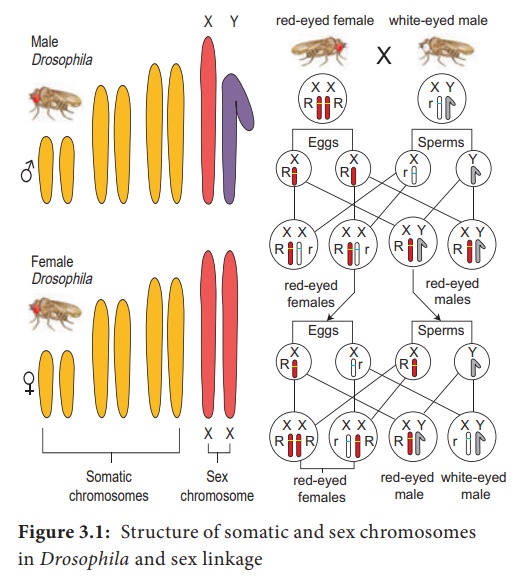
Cette théorie a été largement discutée et controversée par les scientifiques du monde entier. Cependant, ce débat a été définitivement clarifié par les travaux de Thomas Hunt Morgan (1910) sur la mouche des fruits Drosophila melanogaster (2n=8).
Cette mouche des fruits a terminé son cycle de vie en deux semaines. Les allèles pour la couleur des yeux rouges ou blancs sont présents sur le chromosome X mais il n'y a pas de contrepartie pour ce gène sur le chromosome Y.
Ainsi, les femelles ont deux allèles pour ce gène, alors que les mâles n'en ont qu'un (Figure 3.1). Les résultats génétiques étaient entièrement basés sur le comportement méiotique des chromosomes X et Y. De même, les gènes de la couleur jaune du corps et des ailes miniatures sont également portés sur le chromosome X. Cette étude soutient fortement l'idée que les gènes sont situés sur les chromosomes. Les gènes liés reliés entre eux sur le chromosome sexuel sont appelés liens sexuels.
Comparaison entre le comportement des gènes et des chromosomes
Vers le XXe siècle, les cytologistes ont établi que, généralement, le nombre total de chromosomes est constant dans toutes les cellules d'une espèce. Une cellule eucaryote diploïde a deux ensembles haploïdes de chromosomes, un ensemble de chaque parent. Toutes les cellules somatiques d'un organisme portent le même complément génétique. Le comportement des chromosomes pendant la méiose explique non seulement les principes de Mendel, mais conduit à des approches nouvelles et différentes pour étudier l'hérédité.
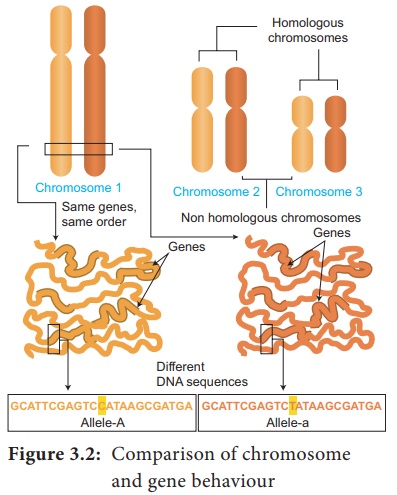
Les aspects importants à retenir sur le comportement des chromosomes lors de la division cellulaire (méiose) sont les suivants.
· Les allèles d'un génotype se retrouvent dans le même locus d'un chromosome homologue (A/a) (Figure 3.2).
· Dans la phase S de l'interphase méiotique, chaque chromosome se réplique en formant deux copies de chaque allèle (AA/aa), une sur chaque chromatide.
· Les chromosomes homologues se séparent en anaphase I, séparant ainsi deux allèles différents (AA) et (aa).
· Dans l'anaphase II de la méiose, la séparation des chromatides sœurs des chromosomes homologues a lieu. Par conséquent, chaque cellule fille (gamète) ne porte qu'un seul allèle (gène) d'un caractère (A), (A), (a) et (a).
Thomas Hunt Morgan (1933) a reçu le prix Nobel de physiologie ou médecine pour ses découvertes concernant le rôle joué par les chromosomes dans l'hérédité.