

La variation génétique entre les individus fournit la matière première de la source ultime des changements évolutifs.

La mutation et la recombinaison sont les deux principaux processus responsables de la variation génétique. Un changement soudain dans le matériel génétique des feuilles mutantes d'un organisme est appelé mutation . Le terme mutation a été introduit par Hugo de Vries (1901) alors qu'il étudiait sur la plante, l'onagre ( Oenothera lamarkiana ) et proposait la « théorie de la mutation » .
Il existe deux grands types de modifications du matériel génétique. Ce sont des mutations ponctuelles et des mutations chromosomiques.
Les événements mutationnels qui se produisent dans des gènes individuels sont appelés mutations géniques ou mutations ponctuelles, tandis que les changements se produisant dans la structure et le nombre de chromosomes sont appelés mutations chromosomiques. Les agents responsables de la mutation sont appelés mutagènes, qui augmentent le taux de mutation. Les mutations peuvent se produire spontanément ou induites. La production de mutants par exposition à des mutagènes est appelée mutagenèse et l'organisme est dit mutagénisé.
1. Types de mutations
Voyons les deux classes générales de mutation génique :
· Les mutations affectant une seule base ou une paire de bases d'ADN sont appelées mutations ponctuelles
· Mutations modifiant le nombre de copies d'une petite séquence nucléotidique répétée au sein d'un gène
Point de mutation
Il fait référence à des altérations de paires de bases uniques d'ADN ou d'un petit nombre de paires de bases adjacentes
Types de mutations ponctuelles
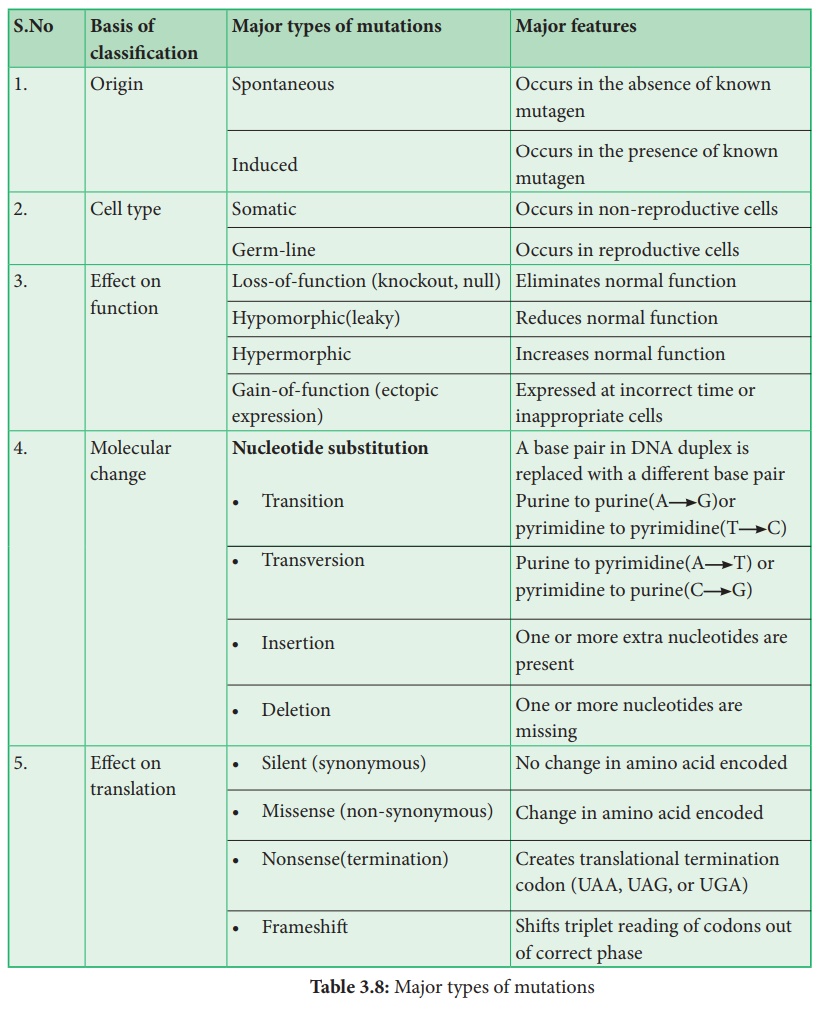
Les mutations ponctuelles dans l'ADN sont classées en deux types principaux. Ce sont des substitutions de paires de bases et des insertions ou suppressions de paires de bases. Les substitutions de bases sont des mutations dans lesquelles il y a un changement dans l'ADN tel qu'une paire de bases est remplacée par une autre (Figure : 3.17). Il peut être divisé en deux sous-types : les transitions et les transversions. Les mutations d'addition ou de délétion sont en fait des additions ou des délétions de paires de nucléotides et également appelées additions ou délétions de paires de bases. Collectivement, elles sont appelées mutations indel ( pour insertion - délétion).
Les mutations de substitution ou les mutations indel affectent la traduction. Sur la base de ces différents types de mutations sont donnés ci-dessous.
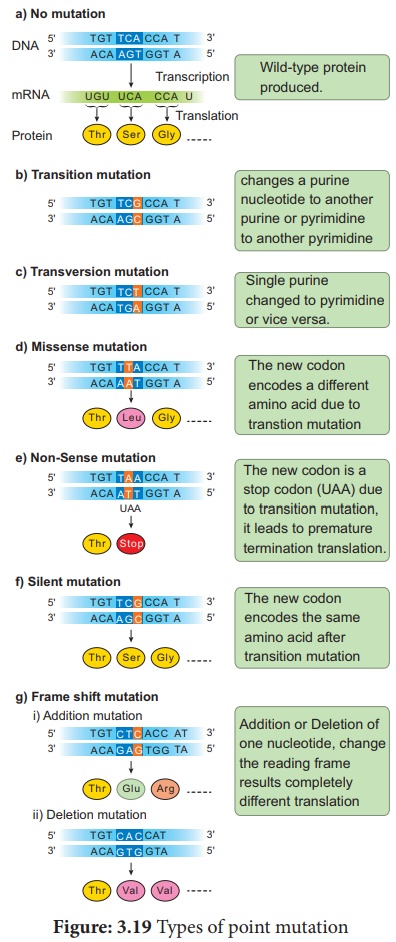
La mutation qui change un codon pour un acide aminé en un autre codon pour ce même acide aminé est appelée mutations synonymes ou silencieuses . La mutation où le codon d'un acide aminé est transformé en un codon d'un autre acide aminé est appelée mutations Missense ou non synonymes . Les mutations où le codon d'un acide aminé est transformé en un codon de terminaison ou d'arrêt sont appelées mutation non-sens . Les mutations qui entraînent l'ajout ou la suppression d'une seule paire de bases d'ADN qui modifie le cadre de lecture du processus de traduction, entraînant une perte complète de la structure et de la fonction normales de la protéine, sont appelées mutations Frameshift . (Figure : 3.19).
2. Agents mutagènes
Les facteurs qui provoquent la mutation génétique sont appelés agents mutagènes ou mutagènes . Les mutagènes sont de deux types, les mutagènes physiques et les mutagènes chimiques. Muller (1927) a été le premier à découvrir un mutagène physique chez la drosophile .
Mutagènes physiques :
Les scientifiques utilisent la température et les radiations telles que les rayons X, les rayons gamma, les rayons alfa, les rayons bêta, les neutrons, les rayons cosmiques, les isotopes radioactifs, les rayons ultraviolets comme mutagène physique pour produire des mutations chez diverses plantes et animaux.
Température : L'augmentation de la température augmente le taux de mutation. Lors de la montée en température, rompt les liaisons hydrogène entre deux nucléotides d'ADN, ce qui affecte le processus de réplication et de transcription.
Rayonnement : le spectre électromagnétique contient des rayons de longueur d'onde plus courte et plus longue que le spectre visible. Ceux-ci sont classés en rayonnements ionisants et non ionisants. Les rayonnements ionisants ont une longueur d'onde courte et transportent une énergie suffisamment élevée pour ioniser les électrons de l'atome. Rayons X, rayons gamma, rayons alfa, rayons bêta et rayons cosmiques qui cassent les chromosomes (mutation chromosomique) et les chromatides dans les cellules irradiées. Rayonnement non ionisant, les rayons UV ont des longueurs d'onde plus longues et transportent une énergie plus faible, ils ont donc un pouvoir de pénétration plus faible que les rayonnements ionisants. Il est utilisé pour traiter les micro-organismes unicellulaires, les spores, les grains de pollen qui possèdent des noyaux situés près de la membrane superficielle.
Sharbati Sonore
Sharbati Sonora est une variété mutante de blé, qui est développée à partir de la variété mexicaine (Sonora 64) par irradiation de rayons gamma. C'est le travail du Dr MSSwaminathan qui est connu comme le « père de la révolution verte indienne » et de son équipe.
Castor Aruna
Castor Aruna est une variété mutante de ricin qui est développée par traitement des graines avec des neutrons thermiques afin d'induire une maturité très précoce (120 jours au lieu de 270 jours comme variété originale).
Mutagènes chimiques :
Les produits chimiques qui induisent une mutation sont appelés mutagènes chimiques. Certains mutagènes chimiques sont le gaz moutarde, l'acide nitreux, les sulfonates de méthane d'éthyle et de méthyle (EMS et MMS), l'uréthane d'éthyle, le sel magnétique, le formaldéhyde, l'éosine et l'enthrosine. Exemple : Le protoxyde d'azote altère les bases azotées de l'ADN et perturbe la réplication et la transcription qui conduisent à la formation de polypeptide incomplet et défectueux lors de la traduction.
Comutagènes
Les composés qui n'ont pas de propriétés mutagènes propres mais qui peuvent renforcer les effets de mutagènes connus sont appelés comutagènes.
Exemple : L'acide ascorbique augmente les dommages causés par le peroxyde d'hydrogène.
La caféine augmente la toxicité du méthotrexate
Gaz moutarde (sulfure de dichloroéthyle) utilisé comme arme chimique pendant la Première Guerre mondiale.
HJ Muller (1928) a utilisé pour la première fois les rayons X pour induire des mutations chez la mouche des fruits.
LJ Stadler a rapporté des mutations induites chez les plantes en utilisant des rayons X et des rayons gamma.
La mutagenèse chimique a été signalée pour la première fois par C. Auerback (1944).
3. Mutations chromosomiques
Le génome peut également être modifié à plus grande échelle en modifiant la structure des chromosomes ou en changeant le nombre de chromosomes dans une cellule. Ces variations à grande échelle sont appelées mutations chromosomiques ou aberrations chromosomiques . Les mutations génétiques sont des modifications qui se produisent au sein d'un gène, tandis que les mutations chromosomiques sont des modifications d'une région chromosomique composée de nombreux gènes. Il peut être détecté par examen microscopique, analyse génétique ou les deux. En revanche, les mutations génétiques ne sont jamais détectables au microscope. Les mutations chromosomiques sont divisées en deux groupes : les modifications du nombre de chromosomes et les modifications de la structure des chromosomes.
1. Modifications du nombre de chromosomes
Chaque cellule d'organismes vivants possède un nombre fixe de chromosomes. Il varie selon les espèces. Même si certaines espèces de plantes et d'animaux ont un nombre identique de chromosomes, leur caractère ne sera pas similaire. Par conséquent, le nombre de chromosomes ne différenciera pas le caractère des espèces les unes des autres, mais la nature du matériel héréditaire (gène) dans le chromosome qui détermine le caractère des espèces.
Parfois, le nombre de chromosomes des cellules somatiques est modifié en raison de l'ajout ou de l'élimination d'un chromosome individuel ou d'un ensemble de base de chromosomes. Cette condition est connue sous le nom d' aberration chromosomique numérique ou de ploïdie . Il existe deux types de ploïdie.
(je). Ploïdie impliquant des chromosomes individuels au sein d'un ensemble diploïde ( aneuploïdie )
(ii). Ploïdie impliquant des ensembles entiers de chromosomes ( Euploïdie ) (Figure 3.20)
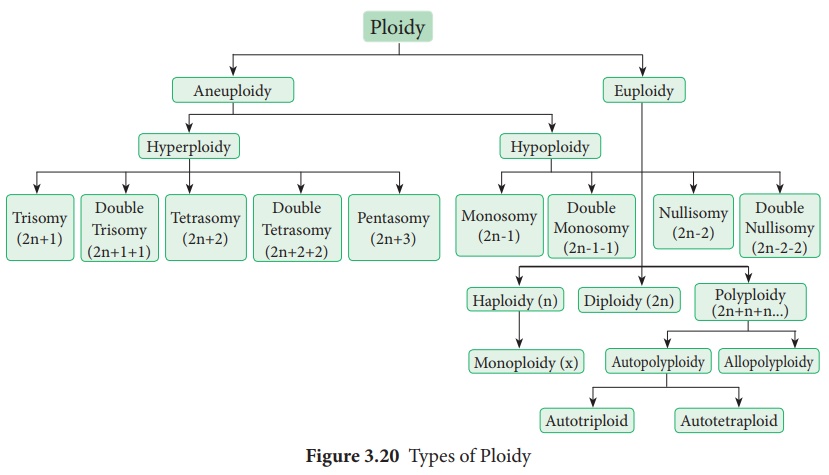
(i) Aneuploïdie
C'est une condition dans laquelle le nombre de diploïdes est modifié soit par l'ajout, soit par la suppression d'un ou de plusieurs chromosomes. Les organismes présentant une aneuploïdie sont appelés aneuploïdes ou hétéroploïdes . Ils sont de deux types, l'hyperploïdie et l'hypoploïdie (Figure 3.21).
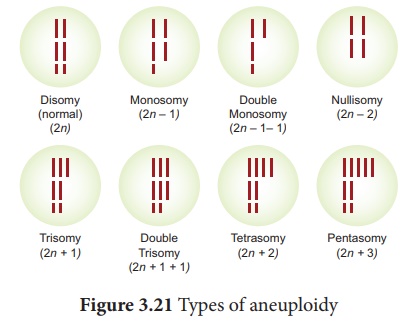
1. Hyperploïdie
L'ajout d'un ou plusieurs chromosomes à des ensembles diploïdes est appelé hyperploïdie . Ensemble diploïde de chromosomes représenté par Disomy. L'hyperploïdie peut être divisée en trois types. Ils sont les suivants,
(a) Trisomie
L'ajout d'un chromosome unique à un ensemble diploïde est appelé trisomie simple (2n + 1). Les trisomiques ont été signalés pour la première fois par Blackeslee (1910) chez Datura stramonium (Jimson weed). Mais plus tard, il a été signalé à Nicotiana, Pisum et Oenothera. Parfois, l'ajout de deux chromosomes individuels de paires chromosomiques différentes à des ensembles diploïdes normaux est appelé double trisomie (2n + 1 + 1).
(b) Tétrasomie
L'ajout d'une paire ou de deux paires individuelles de chromosomes à un ensemble diploïde est appelé respectivement tétrasomie (2n + 2) et double tétrasomie (2n + 2 + 2). Tous les tétrasomiques possibles sont disponibles en blé.
(c) Pentasomie
L'ajout de trois chromosomes individuels de différentes paires chromosomiques à un ensemble diploïde normal est appelé pentasomie (2n + 3).
2. Hypoploïdie
La perte d'un ou plusieurs chromosomes de l'ensemble diploïde dans la cellule est appelée hypoploïdie . Il peut être divisé en deux types. Elles sont
(a) Monosomie
La perte d'un seul chromosome de l'ensemble diploïde est appelée monosomie (2n-1). Cependant, la perte de deux ou trois chromosomes individuels est appelée respectivement double monosomie (2n-1-1) et triple monosomie (2n-1-1-1). Des monosomes doubles sont observés chez le maïs.
(b) Nullisomie
La perte d'une paire de chromosomes homologues ou de deux paires de chromosomes homologues de l'ensemble diploïde est appelée nullisomy (2n-2) et double nullisomy (2n-2-2) respectivement. L'autofécondation des plantes monosomiques produit des nullisomiques. Ils sont généralement mortels.
(ii) Euploïdie
L'euploïdie est une condition dans laquelle les organismes possèdent un ou plusieurs ensembles de base de chromosomes. L'euploïdie est classée en monoploïdie, diploïdie et polyploïdie. La condition dans laquelle un organisme ou une cellule somatique possède deux ensembles de chromosomes est appelée diploïde (2n). La moitié du nombre de chromosomes somatiques est appelée numéro de chromosome gamétique appelé haploïde (n). Il convient de noter que l'haploïdie (n) est différente d'une monoploïdie (x). Par exemple, la plante de blé tendre est une polyploïdie (hexaploïdie) 2n=6x=72 chromosomes. Son nombre haploïde (n) est de 36, mais sa monoploïdie (x) est de 12. Par conséquent, la condition haploïde et diploïde est venue régulièrement l'une après l'autre et le même nombre de chromosomes est maintenu de génération en génération, mais la condition de monoploïdie se produit lorsqu'un organisme est en état de polyploïdie. Dans un vrai diploïde, le nombre de chromosomes monoploïdes et haploïdes est le même. Ainsi, un monoploïde peut être un haploïde mais tous les haploïdes ne peuvent pas être un monoploïde.
Polyploïdie
La polyploïdie est la condition dans laquelle un organisme possède plus de deux ensembles de base de chromosomes. Lorsqu'il y a trois, quatre, cinq ou six ensembles de base de chromosomes, on les appelle respectivement triploïdie (3x), tétraploïdie (4x), pentaploïdie (5x) et hexaploïdie (6x). Généralement, la polyploïdie est très fréquente chez les plantes mais plus rare chez les animaux. Une augmentation du nombre d'ensembles de chromosomes a été un facteur important dans l'origine de nouvelles espèces végétales. Mais un niveau de ploïdie plus élevé conduit à la mort. La polyploïdie est de deux types. Ce sont l'autopolyploïdie et l'allopolyploïdie
1. Autopolyploïdie
L'organisme qui possède plus de deux ensembles haploïdes de chromosomes issus de la même espèce est appelé autopolyploïde. Ils sont divisés en deux types. Autotriploïdes et autotétraploïdes.
Les autotriploïdes ont trois ensembles de leurs propres génomes. Ils peuvent être produits artificiellement par croisement entre des espèces autotétraploïdes et diploïdes. Ils sont hautement stériles en raison de la formation défectueuse de gamètes. Exemple : Les bananiers cultivés sont généralement triploïdes et sont sans pépins et ont des fruits plus gros que les diploïdes. Les betteraves à sucre triploïdes ont une teneur en sucre plus élevée que les diploïdes et sont résistantes aux moisissures. Le doob commun ( Cyanodon dactylon ) est un autotriploïde naturel. La pastèque sans pépins, la pomme, la betterave à sucre, la tomate, la banane sont des autotriploïdes artificiels.
Les autotétraploïdes possèdent quatre copies de leur propre génome. Ils peuvent être induits en doublant les chromosomes d'une espèce diploïde. Exemple : seigle, raisin, luzerne, arachide, pomme de terre et café.
2. Allopolyploïdie
Un organisme qui possède au moins deux ensembles de base de chromosomes dérivés de deux espèces différentes est appelé allopolyploïdie. Il peut être développé par croisements interspécifiques et la fertilité est restaurée par doublement chromosomique avec un traitement à la colchicine. Les allopolyploïdes se forment uniquement entre des espèces étroitement apparentées. (Figure 3.22)
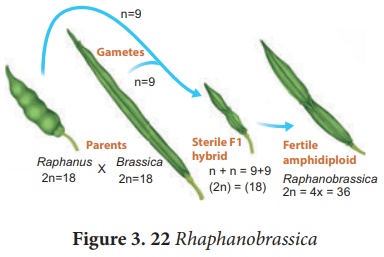
Exemple : 1 Raphanobrassica , GD Karpechenko (1927) un généticien russe, a croisé le radis ( Raphanus sativus , 2n=18) et le chou ( Brassica oleracea , 2n=18) pour produire un hybride F 1 qui était stérile. Quand il a doublé le chromosome de l'hybride F 1 , il l'a rendu fertile. Il s'attendait à ce que cette plante présente la racine de radis et les feuilles comme du chou, ce qui rendrait toute la plante comestible, mais c'était l'inverse, il a donc été très déçu.
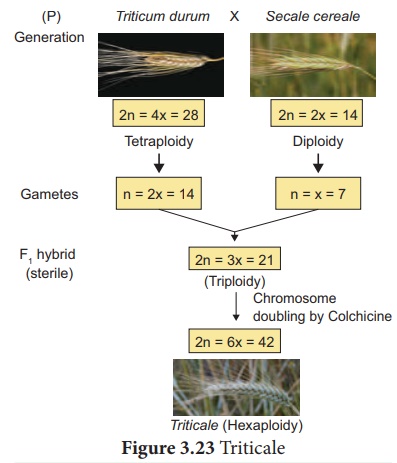
Exemple : 2 Triticale , la première céréale fabriquée par l'homme. Selon le niveau de ploïdie, le triticale peut être divisé en trois groupes principaux.
(je). Tétraploïdie : Croisements entre le blé diploïde et le seigle.
(ii). Hexaploïdie : croisements entre le blé tétraploïde Triticum durum (blé macaroni) et le seigle
(iii). Octoploïdie : croisements entre le blé hexaploïde T. aestivum (blé panifiable) et le seigle
Les plantes hybrides Hexaploïdy Triticale présentent les caractéristiques du blé macaroni et du seigle. Par exemple, ils combinent la teneur élevée en protéines du blé avec la teneur élevée du seigle en acide aminé lysine, qui est faible dans le blé. Cela peut être expliqué par le graphique ci-dessous (Figure : 3.23).
La colchicine , un alcaloïde est extrait de la racine et des bulbes de Colchicum autumnale , lorsqu'elle est appliquée à faible concentration sur les pointes de croissance des plantes, elle induira une polyploïdie. Étonnamment, il n'affecte pas la plante source Colchicum , en raison de la présence d'anticolochicine.

Signification de la ploïdie
· De nombreux polyploïdes sont plus vigoureux et plus adaptables que les diploïdes.
· De nombreuses plantes ornementales sont autotétraploïdes et ont des fleurs plus grosses et une durée de floraison plus longue que les diploïdes.
· Les autopolyploïdes ont généralement une augmentation du poids frais en raison d'une plus grande teneur en eau.
· Les aneuploïdes sont utiles pour déterminer les effets phénotypiques de la perte ou du gain de différents chromosomes.
· De nombreux angiospermes sont allopolyploïdes et jouent un rôle dans l'évolution des plantes.
II Modifications structurelles du chromosome (aberration chromosomique structurelle)
Les variations structurelles causées par l'ajout ou la suppression d'une partie du chromosome entraînant un réarrangement des gènes sont appelées aberrations chromosomiques structurelles . Il se produit en raison de rayonnements ionisants ou de composés chimiques. Sur la base des cassures et des retrouvailles dans les chromosomes, il existe quatre types d'aberrations. Ils sont classés en deux groupes.
A. Changements dans le nombre de locus de gènes
1. Suppression ou lacune
2. Duplication ou répétition
B. Changements dans l'arrangement des locus de gènes
3. Renversement
4. Translocation
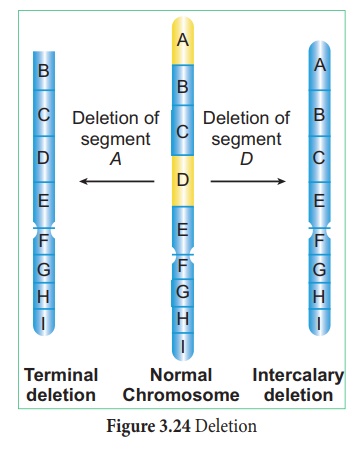
1. Suppression ou lacune
La perte d'une portion de chromosome est appelée délétion. Sur la base de l'emplacement de la rupture sur le chromosome, il est divisé en délétion terminale et délétion intercalaire. Il se produit en raison de produits chimiques, de médicaments et de radiations. On l'observe chez la Drosophile et le Maïs. (Figure 3.24)
Il existe deux types de suppression :
je. Délétion terminale : rupture unique à l'une des extrémités du chromosome.
ii. Délétion intercalaire ou délétion interstitielle : Elle est causée par deux cassures et réunion des parties terminales partant du milieu.
Les deux délétions sont observables au cours du stade pachytène méiotique et du chromosome polytène. La boucle non appariée formée dans la partie chromosomique normale au moment de l'appariement chromosomique. Ces boucles sont appelées boucles de déficience et peuvent être observées dans la prophase méiotique. Des délétions plus importantes peuvent entraîner un effet létal.
2. Duplication ou répétition
Le processus d'arrangement du même ordre de gènes répété plus d'une fois dans le même
En raison de la duplication, certains gènes sont présents en plus de deux copies. Il a été signalé pour la première fois chez Drosophila par Bridges (1919) et d'autres exemples sont le maïs et le pois . C'est trois types.
je. Duplication en tandem
Le segment dupliqué est situé immédiatement après le segment normal du chromosome dans le même ordre.
ii. Duplication en tandem inversée
Le segment dupliqué est situé immédiatement après le segment normal mais l'ordre de la séquence du gène sera inversé.
iii. Duplication déplacée
Le segment dupliqué est situé sur le même chromosome, mais loin du segment normal. (Figure 3.25)
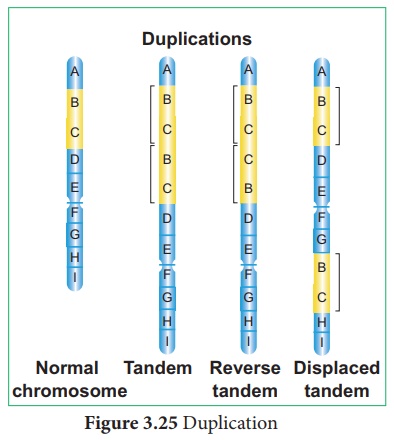
Les duplications jouent un rôle majeur dans l'évolution.
3. Renversement
Un réarrangement de l'ordre des gènes dans un chromosome par inversion d'un angle de 180°. Cela implique deux cassures chromosomiques et une réunion. Au cours de ce processus, il n'y a ni gain ni perte, mais les séquences de gènes sont réarrangées. L'inversion a été signalée pour la première fois chez la drosophile par Sturtevant (1926). Il existe deux types d'inversion, paracentrique et péricentrique (Figure 3.26).
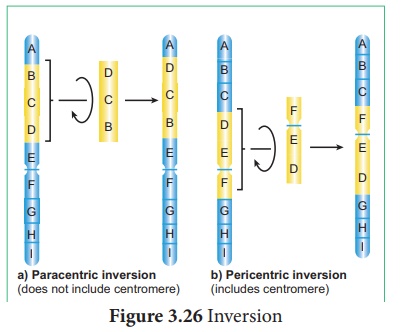
je. Inversion paracentrique : Une inversion qui a lieu en dehors du centromère
ii. Inversion péricentrique : Une inversion qui inclut le centromère.
Les inversions conduisent à l'évolution d'une nouvelle espèce.
4. Translocation
Le transfert d'un segment de chromosome à un chromosome non homologue est appelé translocation. La translocation ne doit pas être confondue avec le croisement, dans lequel un échange de matériel génétique entre chromosomes homologues a lieu. La translocation se produit à la suite d'un échange de segments chromosomiques dans des chromosomes non homologues. Il existe trois types
je. Translocation simple
ii. Décalage de la translocation
iii. Translocation réciproque
je. Translocation simple
Une seule cassure est faite dans un seul chromosome. Le segment cassé s'attache à une extrémité d'un chromosome non homologue. Il se produit très rarement dans la nature.
ii. Décalage de la translocation
Un segment brisé d'un chromosome est inséré de manière interstitielle dans un chromosome non homologue.
iii. Translocations réciproques
Il s'agit d'un échange mutuel de segments chromosomiques entre deux chromosomes non homologues . On l'appelle aussi croisement illégitime . Il est ensuite divisé en deux types (Figure 3.27).
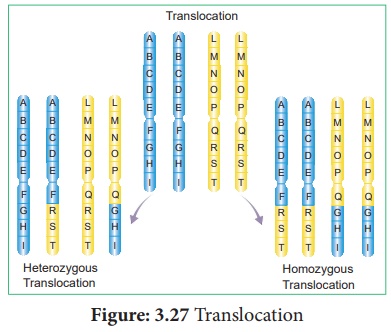
un. Translocation homozygote : Les deux chromosomes de deux paires sont impliqués dans la translocation. Deux homologues de chaque chromosome transloqué sont identiques.
b. Translocation hétérozygote : Un seul des chromosomes de chaque paire de deux homologues est impliqué dans la translocation, tandis que le chromosome restant est normal.
Les translocations jouent un rôle majeur dans la formation des espèces.