

Le croisement est un processus biologique qui produit une nouvelle combinaison de gènes en interchangeant les segments correspondants entre les chromatides non sœurs d'une paire homologue de chromosomes. Le terme «crossing over» a été inventé par Morgan (1912). Elle a lieu au stade pachytène de la prophase I de la méiose. Habituellement, le croisement se produit dans les cellules germinales pendant la gamétogenèse. C'est ce qu'on appelle le croisement méiotique ou germinal. Il a une occurrence universelle et a une grande signification. Rarement, le croisement se produit dans les cellules somatiques pendant la mitose. On l'appelle croisement somatique ou mitotique.
1. Mécanisme de croisement
Le croisement est un processus précis qui comprend des étapes telles que la synapse, la formation de tétrades, le croisement et la terminalisation.
(i) Synapse
L'appariement intime entre deux chromosomes homologues est initié au stade zygotène de la prophase I de la méiose I. Les chromosomes homologues sont alignés côte à côte, ce qui donne une paire de chromosomes homologues appelés bivalents . Ce phénomène d'appariement est appelé synapsis ou syndesis. Il est de trois types,
1. Synapse procentrique : L'appariement commence au milieu du chromosome.
2. Synapsis proterminal : L'appariement commence à partir des télomères.
3. Synapsis aléatoire : l' appariement peut commencer de n'importe où.
(ii) Formation de tétrades
Chaque chromosome homologue d'un bivalent commence à former deux chromatides sœurs identiques, qui restent maintenues ensemble par un centromère. A ce stade, chaque bivalent a quatre chromatides. Cette étape est appelée étape tétrade .
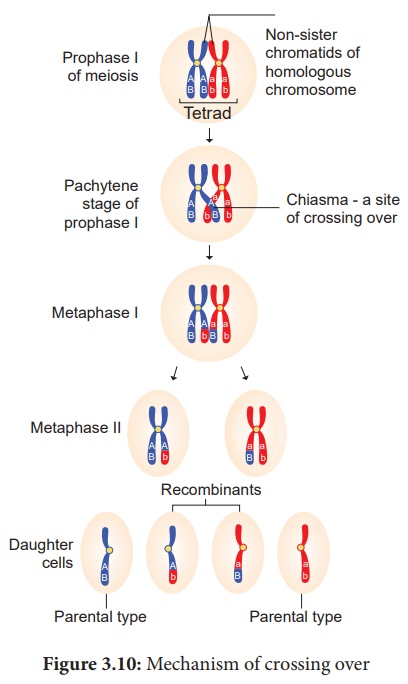
(iii) Croisement
Après la formation des tétrades, le croisement se produit au stade pachytène. Les chromatides non sœurs d'une paire homologue établissent un contact en un ou plusieurs points. Ces points de contact entre chromatides non soeurs de chromosomes homologues sont appelés Chiasmata (singulier-Chiasma). Au chiasma, des structures en forme de croix ou en forme de X se forment, où se produisent la rupture et la réunion de deux chromatides. Il en résulte un échange réciproque de segments égaux et correspondants entre eux. Une étude récente révèle que la formation des synapses et des chiasmas est facilitée par une structure hautement organisée de filaments appelée Synaptonemal Complex (SC) (Figure 3.9). Cette formation de complexe synaptonémique est absente chez certaines espèces de drosophile mâlepar conséquent, le croisement n'a pas lieu.
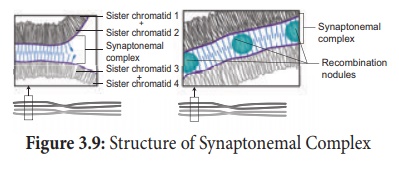
(iv) Terminalisation
Après avoir traversé, le chiasma commence à se déplacer vers l'extrémité terminale des chromatides. C'est ce qu'on appelle la terminalisation . En conséquence, une séparation complète des chromosomes homologues se produit. (Figure 4.10)
2. Types de traversée
Selon le nombre de chiasmes formés, le croisement peut être classé en trois types. (Figure 3.11)
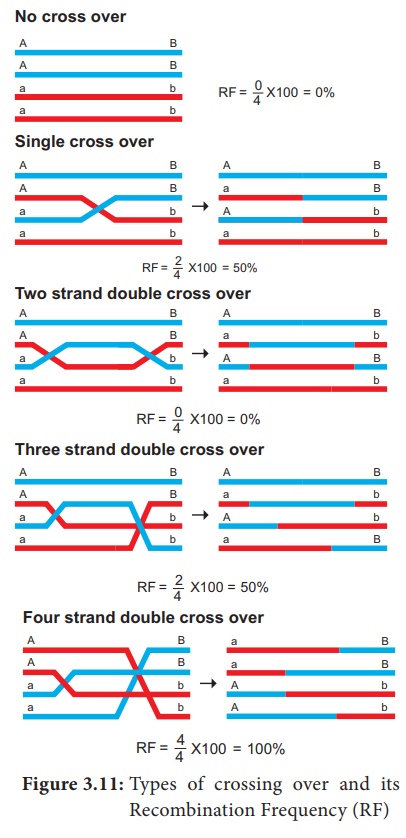
1. Single cross over : Formation d'un seul chiasma et n'impliquant que deux chromatides sur quatre.
2. Double croisement : Formation de deux chiasmes et implique deux ou trois ou les quatre brins
3. Multiple cross over : La formation de plus de deux chiasmes et la fréquence de croisement sont extrêmement faibles.
3. Importance de traverser
Le croisement se produit dans tous les organismes comme les bactéries, les levures, les champignons, les plantes supérieures et les animaux. Son importance est
· L'échange de segments conduit à de nouvelles combinaisons de gènes qui jouent un rôle important dans l'évolution.
· Des études de croisement révèlent que les gènes sont disposés linéairement sur les chromosomes.
· Les cartes génétiques sont établies en fonction de la fréquence des croisements.
· Crossing over aide à comprendre la nature et le mécanisme d'action des gènes.
· Si une nouvelle combinaison utile est formée, elle peut être utilisée dans la sélection végétale.
4. Recombinaison
Le croisement aboutit à la formation d'une nouvelle combinaison de caractères dans un organisme appelé recombinants. En cela, des segments d'ADN sont brisés et recombinés pour produire de nouvelles combinaisons d'allèles. Ce processus est appelé recombinaison. (Figure 3.12)
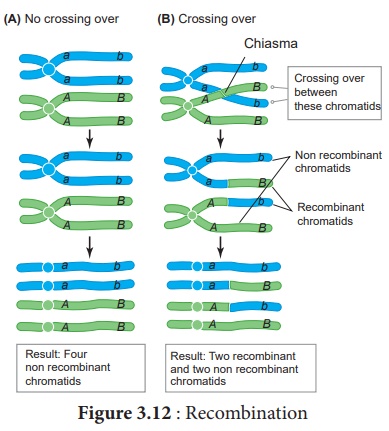
Le modèle largement accepté de recombinaison de l'ADN lors du croisement est le modèle d'ADN hybride de Holliday . Il a été proposé pour la première fois par Robin Holliday en 1964. Il comporte plusieurs étapes. (Figure 3.13)
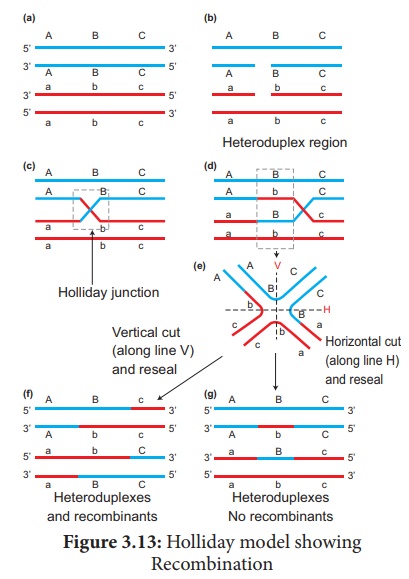
1. Les molécules d'ADN homologues sont appariées côte à côte avec leurs copies dupliquées d'ADN
2. Un brin des deux ADN coupé en un seul endroit par l'enzyme endonucléase .
3. Les brins coupés se croisent et rejoignent les brins homologues formant la structure Holliday ou jonction Holliday .
4. La jonction Holliday migre loin du site d'origine, un processus appelé migration de branche, à la suite de quoi une région hétéroduplex est formée.
5. Les brins d'ADN peuvent couper le long de la ligne verticale (V) ou de la ligne horizontale (H).
6. La coupe verticale se traduira par des hétéroduplex avec des recombinants.
7. La coupe horizontale entraînera un hétéroduplex avec des non recombinants.
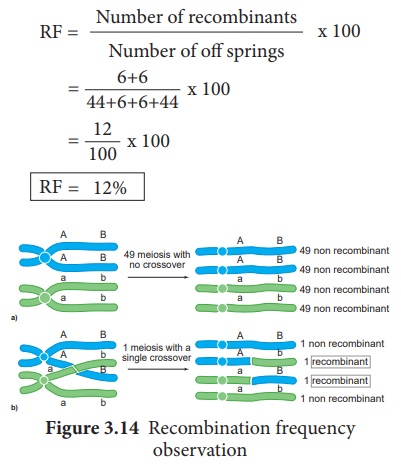
Calcul de la fréquence de recombinaison (RF)
Le pourcentage de descendants recombinants dans un croisement est appelé fréquence de recombinaison. La fréquence de recombinaison (fréquence de croisement) (RF) est calculée à l'aide de la formule suivante. Les données sont obtenues à partir d'allèles en configuration de couplage (Figure 3.14)
5. Cartographie génétique
Les gènes sont présents dans un ordre linéaire le long du chromosome. Ils sont présents dans un endroit précis appelé locus (pluriel : loci). La représentation schématique de la position des gènes et des distances associées entre les gènes adjacents est appelée cartographie génétique . Elle est directement proportionnelle à la fréquence de recombinaison entre eux. Elle est également appelée carte de liaison . Le concept de cartographie génétique a été développé pour la première fois par l'étudiant de Morgan, Alfred H Sturtevant , en 1913. Il fournit des indices sur l'emplacement des gènes sur ce chromosome.
Distance de la carte
L'unité de distance dans une carte génétique est appelée une unité de carte (mu). Une unité cartographique équivaut à un pour cent de franchissement (figure 4. ). Une unité cartographique est également appelée centimorgan (cM) en l'honneur de TH Morgan . 100 centimorgan est égal à un Morgan (M). Par exemple : Une distance entre les gènes A et B est estimée à 3,5 unités cartographiques. Elle est égale à 3,5 centimorgans soit 3,5 % soit 0,035 fréquence de recombinaison entre les gènes.
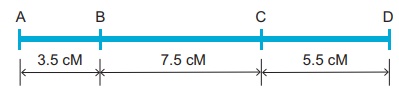
Les cartes génétiques peuvent être construites à partir d'une série de croisements tests pour des paires de gènes appelés croisements à deux points . Mais ce n'est pas efficace car le double croisement est manqué.
Croix de test à trois points
Une technique de cartographie plus efficace consiste à construire sur la base des résultats du test croisé à trois points. Il se réfère à l'analyse des modèles d'hérédité de trois allèles par test croisant un hétérozygote triple récessif avec un homozygote triple récessif. Il permet de déterminer la distance entre les trois allèles et l'ordre dans lequel ils se situent sur le chromosome. Les doubles croisements peuvent être détectés, ce qui fournira des distances cartographiques plus précises.
La croix de test à trois points peut être mieux comprise en considérant un exemple.
Dans le maïs (maïs), les trois allèles récessifs sont
1. l pour une croissance paresseuse ou prostrée
2. g pour feuille brillante
3. s pour endosperme sucré
Ces trois allèles récessifs (lgs) sont croisés avec des allèles dominants de type sauvage (LGS).

Parents LGS / LGS x lgs / lgs
Gamètes LGS x lgs
F1 trihybride LGS / lgs
Croix d'essai
(Croisements hétérozygotes F1 avec
allèles triples récessifs)
LGS / lgs x lgs / lgs
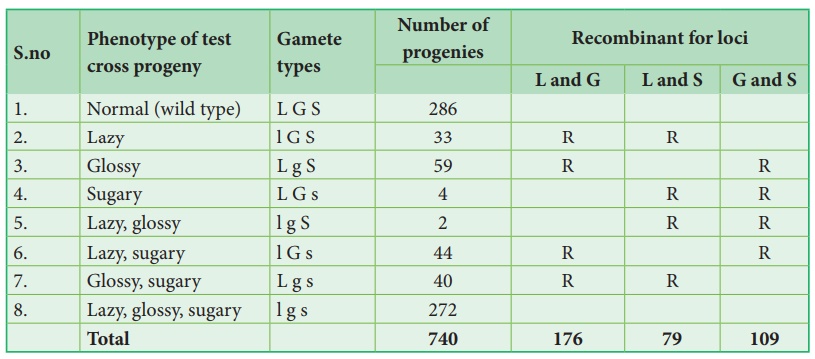
Ce croisement test trihybride produit 8 types différents (23=8) de gamètes dans lesquels 740 descendances sont observées. Le tableau suivant montre le résultat obtenu à partir d'un croisement test de maïs avec trois gènes liés.
L'analyse d'une croix à trois points:
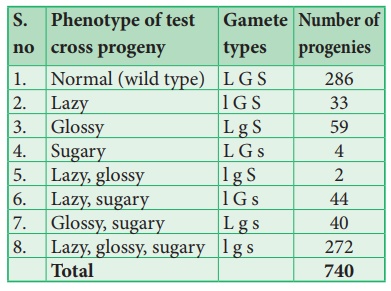
À partir du résultat ci-dessus, il faut faire attention à observer les types parentaux (P) et recombinants (R). Notez d'abord que les génotypes parentaux pour les triples homozygotes sont LGS et lgs, puis analysez deux locus recombinants à la fois ordonnés LG/ lg, LS/ ls et GS/ g s. En cela, toute combinaison autre que ces deux constitue un recombinant (R).
Analysons les locus de deux allèles à la fois en commençant par L et G Puisque les génotypes parentaux LG et lg, les recombinants seront L g et l G. La fréquence de recombinaison (RF) pour ces deux allèles peut être calculée comme suit

Pour les loci L et S, les recombinants sont L s et l S. La fréquence de recombinaison (RF) sera la suivante
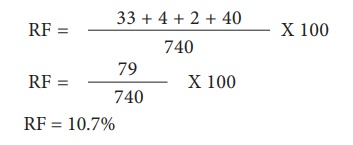
Pour les loci G et S, les recombinants sont G s et g S. La fréquence de recombinaison (RF) sera la suivante
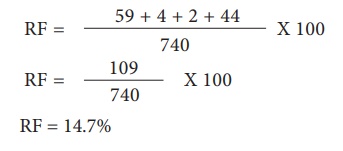
Tous les locus sont liés, car toutes les valeurs RF sont largement inférieures à 50 %. Dans ce LG, les locus affichent la valeur RF la plus élevée, ils doivent être les plus éloignés. Par conséquent, le lieu S doit se situer entre eux. L'ordre des gènes devrait être ls g. Une carte génétique peut être tracée comme suit : (Figure 3.15)

Un dernier point notez que deux distances cartographiques plus petites, 10,7 mu et 14,7., s'additionnent pour donner 25,4 mu, ce qui est supérieur à 23,7 mu, la distance calculée pour l et g. nous devons identifier les deux plus petits nombres de descendants (au total 8) par rapport à la recombinaison de L et G. Ces deux plus petits descendants sont des doubles recombinants issus d'un double croisement. Les deux descendances les moins comptées non seulement une fois auraient dû compter chacune deux fois car chacune représente une double descendance recombinante.
Par conséquent, nous pouvons corriger la valeur en ajoutant les nombres 33+59+44+40+4+4+2+2=188. Sur le total de 740, ce nombre exactement 25,4 %, ce qui est identique à la somme de deux valeurs composantes.
Le test de combinaison parentale croisée peut être réécrit comme suit :
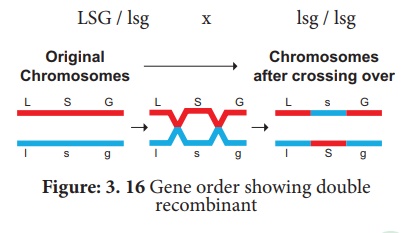
Utilisations de la cartographie génétique
· Il est utilisé pour déterminer l'ordre des gènes, identifier le locus d'un gène et calculer les distances entre les gènes.
· Ils sont utiles pour prédire les résultats des croisements dihybrides et trihybrides.
· Il permet aux généticiens de comprendre la complexité génétique globale d'un organisme particulier.