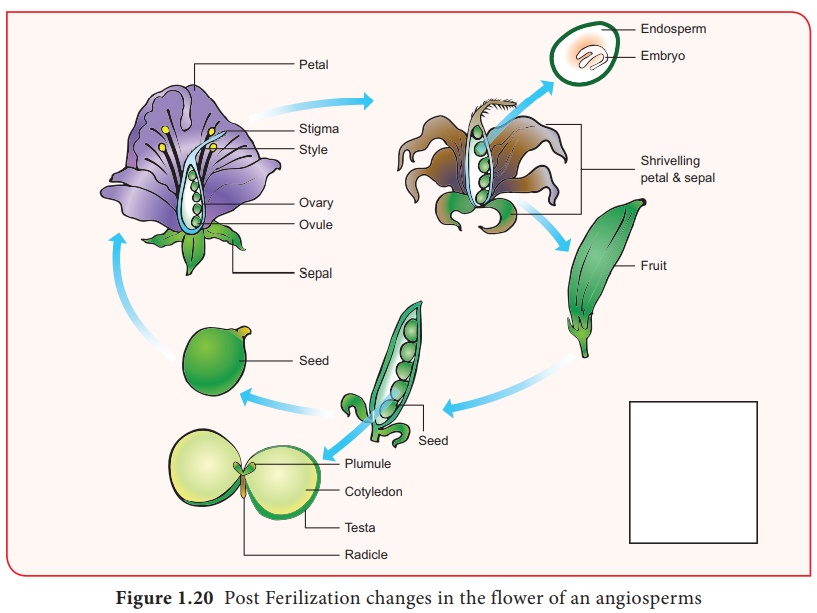
Après la fécondation, plusieurs changements ont lieu dans les parties florales jusqu'à la formation de la graine (Figure 1.20).
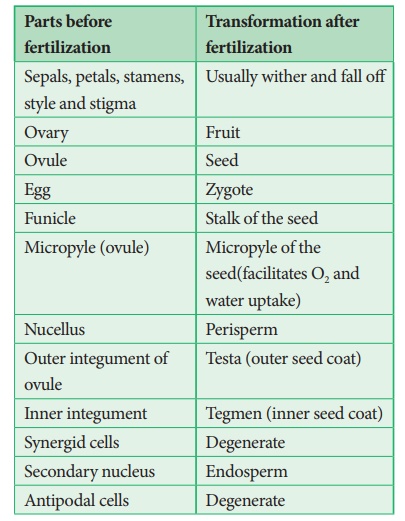
Les événements après la fécondation (endosperme, développement embryonnaire, formation de graines, fruits) sont appelés changements post-fécondation.
Endosperme
Le noyau primaire de l'endosperme (PEN) se divise immédiatement après la fécondation, mais avant que le zygote ne commence à se diviser, en un endosperme. Le noyau primaire de l'endosperme est le résultat d'une triple fusion (deux noyaux polaires et un noyau de spermatozoïde) et possède donc un nombre de chromosomes 3n. C'est un tissu nutritif et une structure régulatrice qui nourrit l'embryon en développement. Selon le mode de développement, trois types d'endosperme sont reconnus dans les angiospermes. Il s'agit de l'albumen nucléaire, de l'endosperme cellulaire et de l'endosperme héliobial (Figure 1.21).
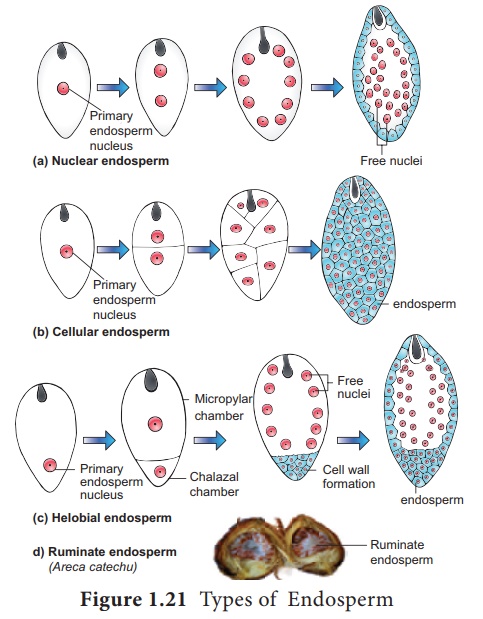
Endosperme nucléaire : Dans le type nucléaire, le noyau primaire de l'endosperme (PEN) se divise en deux sans aucune formation de paroi. La division ultérieure de ces deux noyaux est nucléaire libre de sorte que l'endosperme se compose uniquement de noyaux libres et de cytoplasme autour d'eux. Les noyaux peuvent soit rester libres, soit se séparer par des parois à des stades ultérieurs. Exemples : Coccinia, Capsella et Arachis.
Endosperme cellulaire : Dans ce type, le noyau primaire de l' endosperme (PEN) se divise en 2 noyaux qui sont immédiatement suivis d'une formation de paroi. Les divisions suivantes sont également suivies de murs. Exemples : Adoxa , Helianthus et Scoparia .
Endosperme hélobie: Dans le type hélobie, le noyau primaire de l'endosperme (PEN) se déplace vers la base du sac embryonnaire où il se divise en deux noyaux. Ces 2 noyaux sont séparés par une paroi pour former une grande chambre micropylaire et une petite chambre chalazale. Le noyau de la chambre micropylaire subit plusieurs divisions nucléaires libres alors que celui de la chambre chalazale peut se diviser ou non. Exemples : Hydrilla et Vallisneria
Les endospermes peuvent soit être complètement consommés par l'embryon en développement, soit persister dans les graines matures. Les graines sans endospermes sont appelées graines non endospermes ou ex-albumineuses. Exemples : Pois, Arachide et Haricots . Ces graines avec des endospermes sont appelées graines endospermes ou albumineuses. Les endospermes de ces graines alimentent l'embryon pendant la germination des graines. Exemples : Paddy, Coconut et Castor .
Endosperme ruminé : L'endosperme dont la surface présente des irrégularités et des irrégularités forme l'endosperme ruminé (Exemple : Areca catechu ). L'activité du tégument ou de l'endosperme lui-même entraîne ce type d'endosperme. L'allongement radial inégal de la couche de tégument entraîne la rumination de l'endosperme chez Passiflora . Chez les Annonaceae et les Aristolochiaceae, la croissance ou le repliement défini du tégument produit un endosperme ruminé. La surface irrégulière du tégument fait ruminer l'endosperme chez Myristica .
Fonctions de l'endosperme :
· C'est le tissu nutritif de l'embryon en développement.
· Dans la majorité des angiospermes, le zygote ne se divise qu'après le développement de l'endosperme.
· L' endosperme régule le mode précis de développement de l'embryon.
Endosperme haustoria
Une autre caractéristique intéressante de l'albumen est la présence d'haustoria. Dans le cas de l'endosperme héliobial, la chambre chalazale elle-même agit comme une structure haustoriale.
Dans l'endosperme cellulaire et nucléaire, des structures spéciales sont produites vers le micropylaire, le chalazal, à la fois micropylaire et chalazal qui peuvent être dans la direction latérale selon les espèces. Ceux-ci absorbent les nutriments d'autres tissus externes ou du tissu ovarien et les fournissent à l'embryon en croissance.
Embryogenèse
Développement de l'embryon de dicotylédone
Le développement de l'embryon de Dicot ( Capsella bursa-pastoris) est de type Onagrad ou crucifère. L'embryon se développe à l'extrémité micropylaire du sac embryonnaire.
Le Zygote se divise par une division transversale formant une cellule supérieure ou terminale et une cellule inférieure ou basale . La cellule basale se divise transversalement et la cellule terminale se divise verticalement pour former un proembryon à quatre cellules. Une deuxième division verticale à angle droit par rapport à la première a lieu dans la cellule terminale formant un étage à quatre cellules appelé quadrant. Une division transversale dans le quadrant donne huit cellules disposées en deux niveaux de quatre, chacun appelé étage octant.
Le niveau supérieur de quatre cellules de l'octant est appelé octant épibasal ou antérieur et le niveau inférieur de quatre cellules constitue l' octant hypobasal ou postérieur. Une division périclinale dans les octants entraîne la formation d'un stade à 16 cellules avec huit cellules à l'extérieur et huit à l'intérieur.
Les huit cellules externes représentent le dermatogène et subissent une division anticlinale pour produire l'épiderme. Les huit cellules internes se divisent par division verticale et transversale pour former une couche externe de periblem qui donne naissance au cortex et à une région centrale de pleurome qui forme une stèle
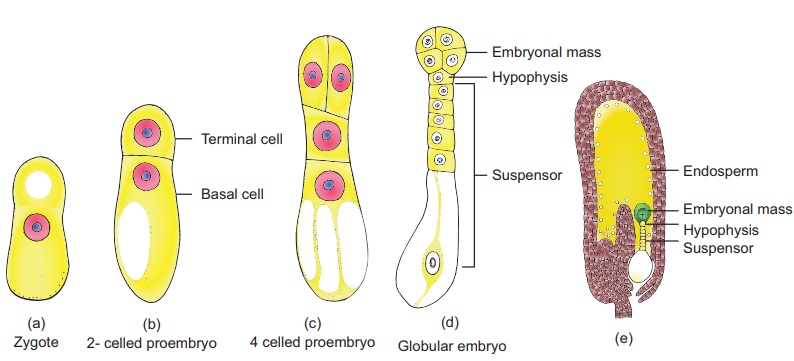
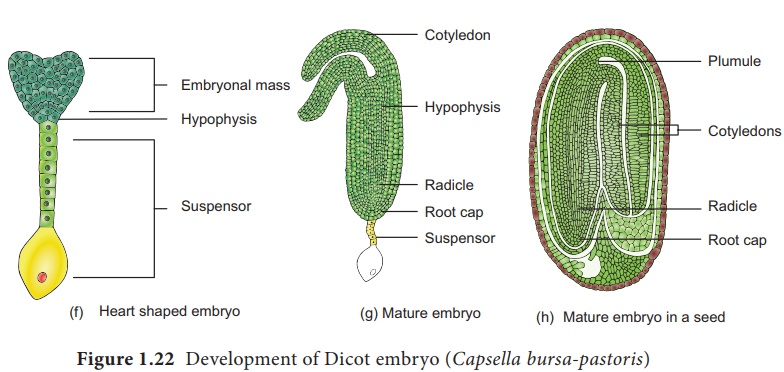
Au cours du développement, les deux cellules de la cellule basale subissent plusieurs divisions transversales pour former un suspenseur de six à dix cellules . L'embryon à ce stade devient globuleux et le suspenseur aide à pousser l'embryon profondément dans l'endosperme. La cellule la plus haute du suspenseur s'agrandit pour former un haustorium . La cellule la plus basse du suspenseur est appelée hypophyse . Une division transversale et deux divisions verticales à angle droit l'une par rapport à l'autre de l'hypophyse entraînent la formation de huit cellules. Les huit cellules sont disposées en deux niveaux de quatre cellules chacun. Le niveau supérieur donne naissance à la coiffe et à l'épiderme. À ce stade, l'embryon proprement dit apparaît en forme de cœur, les divisions cellulaires dans l' hypocotyleet les régions des cotylédons de l'embryon proprement dit entraînent un allongement. Le développement ultérieur donne un embryon en forme de fer à cheval incurvé dans le sac embryonnaire. L'embryon mature a une radicule, deux cotylédons et une plumule (Figure 1.22).
Planter
L'ovule fécondé est appelé graine et possède un embryon, un endosperme et un manteau protecteur. Les graines peuvent être endospermes (blé, maïs, orge et tournesol) ou non endospermes. (Haricot, Mangue, Orchidées et cucurbitacées).
Structure d'une graine Cicer comme exemple pour la graine Dicot
Les graines mûres sont attachées à la paroi du fruit par une tige appelée funicule . Le funicule disparaît en laissant une cicatrice appelée hile . Sous le hile, un petit pore appelé micropyle est présent. Il facilite l'entrée d'oxygène et d'eau dans les graines pendant la germination. Chaque graine a un revêtement extérieur épais appelé tégument. Le tégument est développé à partir des téguments de l'ovule. La couche externe s'appelle testa et est dure tandis que la couche interne est mince, membraneuse et s'appelle tegmen. Dans la plante de pois, le tegmen et le testa sont fusionnés. Deux cotylédons attachés latéralement à l'axe embryonnaire sont présents. Il stocke les matières alimentaires dans le pois alors que dans d'autres graines comme le ricin, l'endosperme contient de la nourriture de réserve et les cotylédons sont minces. La partie embryonnaire
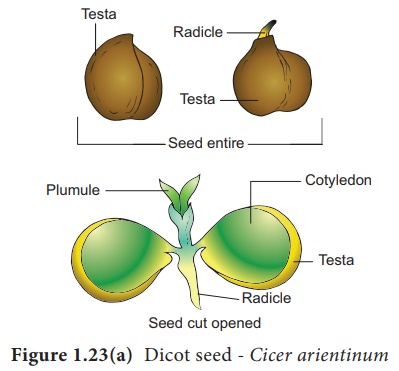
L'autre extrémité de l'axe appelée pousse embryonnaire est la plumule . L'axe embryonnaire au-dessus du niveau du cotylédon est appelé épicotyle tandis que la région cylindrique entre le niveau du cotylédon est appelée hypocotyle (Figure 1.23 a). L'épicotyle se termine par une plumule tandis que l'hypocotyle se termine par une radicule.
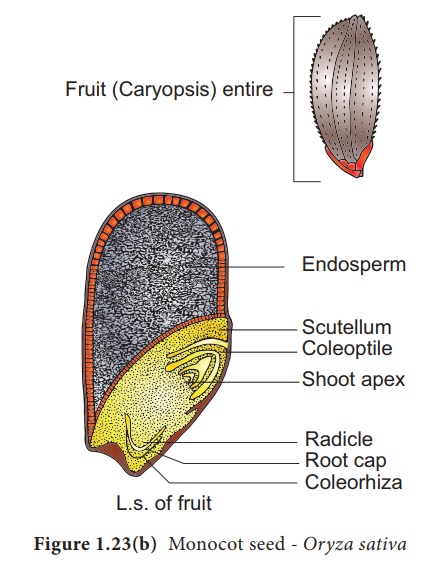
Structure de la graine Oryza comme exemple pour la graine Monocot
La graine de paddy est une graine et s'appelle Caryopsis . Chaque graine reste entourée d' une enveloppe brunâtre constituée de glumes disposées en deux rangées. Le tégument est une couche membraneuse brunâtre qui adhère étroitement au grain. L'endosperme forme la majeure partie du grain et constitue le tissu de stockage. Il est séparé de l'embryon par une couche définie appelée épithélium . L'embryon est petit et se compose d'un cotylédon en forme de bouclier appelé scutellum présent vers le côté latéral de l'axe embryonnaire. Un axe court avec plumule et radicule protégés par la coiffe racinaire est présent. La plumule est entourée d'une gaine protectrice appelée coléoptile. La radicule comprenant la coiffe racinaire est également recouverte d'une gaine protectrice appelée coleorhiza . Le scutellum fournit à l'embryon en croissance le matériel alimentaire absorbé par l'endosperme à l'aide de l'épithélium (Figure 1.23 b).