Structure et événements de pré-fertilisation
Les changements hormonaux et structurels de la plante conduisent à la différenciation et au développement du primordium floral. Les structures et événements impliqués dans la pré-fécondation sont donnés ci-dessous
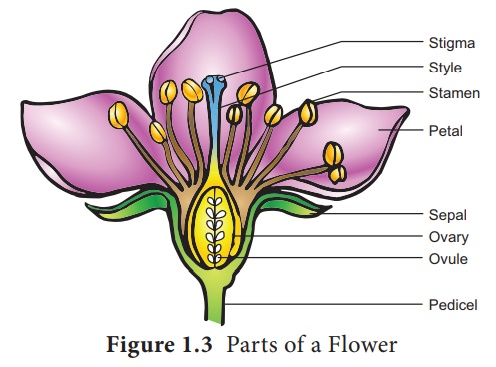
Partie reproductrice masculine - Androcée
L'androcée est constituée d'étamines. Chaque étamine possède une anthère et un filament. L'anthère porte des grains de pollen qui représentent le gamétophyte mâle. Dans ce chapitre, nous discuterons en détail de la structure et du développement de l'anthère.
Développement de l'anthère :
Une très jeune anthère se développe sous la forme d'une masse homogène de cellules entourées d'un épiderme. Au cours de son développement, l'anthère prend une structure à quatre lobes. Dans chaque lobe, une rangée ou quelques rangées de cellules hypodermiques s'agrandissent avec des noyaux bien visibles. Cela fonctionne comme archesporium. Les cellules archésporiales se divisent par divisions périclinales pour former des cellules pariétales primaires vers l'épiderme et des cellules sporogènes primaires vers la face interne de l'anthère. Les cellules pariétales primaires subissent une série de divisions périclinales et anticlinales et forment 2 à 5 couches de parois d'anthères composées d'endothécie, de couches moyennes et de tapetum, de la périphérie au centre.
Microsporogenèse: Les étapes impliquées dans la formation de microspores haploïdes à partir de la cellule mère de microspores diploïdes via la méiose sont appelées microsporogenèse . Les cellules sporogènes primaires directement, ou peuvent subir quelques divisions mitotiques pour former un tissu sporogène .
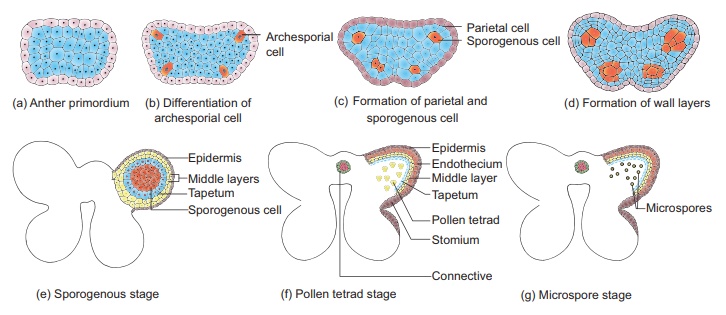
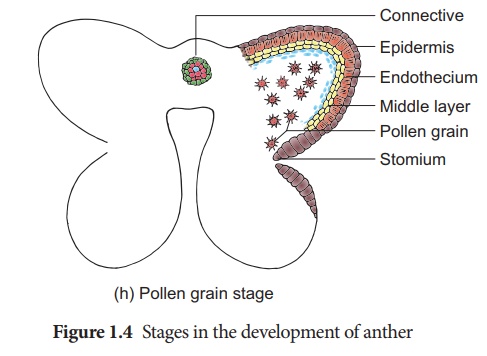
La dernière génération de tissus sporogènes fonctionne comme des cellules mères de microspores. Chaque cellule mère de microspores se divise méiotiquement pour former une tétrade de quatre microspores haploïdes (microspores tétrade). La tétrade de microspores peut être disposée de manière tétraédrique, décussée, linéaire, en forme de T ou isobilatérale. Les microspores se séparent rapidement les unes des autres et restent libres dans la loge de l'anthère et se développent en grains de pollen. Les étapes du développement des microsporanges sont présentées à la figure 1.4. Chez certaines plantes, toutes les microspores d'un microsporange restent réunies, appelées pollinium . Exemple : Calotropis . Les grains de pollen composés se trouvent dans Drosera et Drymis .
TS d'anthère mature
La coupe transversale de l'anthère mature révèle la présence d'une cavité d'anthère entourée d'une paroi d'anthère. Il est bilobé, chaque lobe ayant 2 thèques (dithéque). Une anthère typique est tétrasporangée. Le TS de l'anthère mature est donné à la figure 1.5.
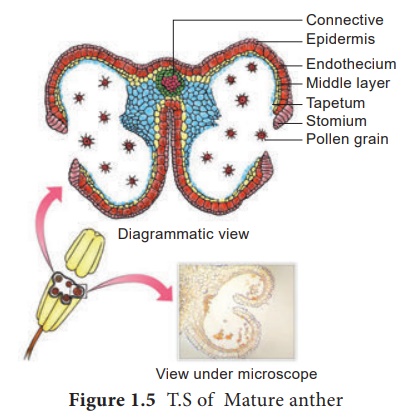
1. Mur d'anthères
La paroi de l'anthère mature se compose des couches suivantes a. Épiderme b. Endothèque c. Couches intermédiaires d. Tapetum.
une. Épiderme : Il est monocouche et protecteur en fonction. Les cellules subissent des divisions anticlinales répétées pour faire face à l'augmentation rapide des tissus internes.
b. Endothèque : Il s'agit généralement d'une seule couche de cellules radialement allongées se trouvant sous l'épiderme. La paroi tangentielle interne développe des bandes (parfois parois radiales également) de cellulose (parfois aussi légèrement lignifiée). Les cellules sont hygroscopiques . Dans les anthères des plantes aquatiques, les saprophytes, les fleurs cléistogames et les parasites extrêmes, la différenciation endothéliale est absente. Les cellules le long de la jonction des deux sporanges d'un lobe d'anthère manquent de ces épaississements. Cette région est appelée stomium . Cette région ainsi que la nature hygroscopique de l'endothécie aident à la déhiscence de l'anthère à maturité.
c. Couches intermédiaires : Deux à trois couches de cellules à côté de l'endothécie constituent les couches intermédiaires. Ils sont généralement éphémères. Ils se désagrègent ou s'écrasent à maturité.
ré. Tapetum : C'est la couche la plus interne de la paroi de l'anthère et atteint son développement maximal au stade tétrade de la microsporogenèse. Il est dérivé en partie de la couche de paroi périphérique et en partie du tissu conjonctif de l'anthère tapissant la loge de l'anthère. Ainsi, le tapetum est d'origine double. Il nourrit le tissu sporogène en développement, les cellules mères des microspores et les microspores. Les cellules du tapetum peuvent rester uninucléées ou peuvent contenir plus d'un noyau ou le noyau peut devenir polyploïde. Il contribue également aux matériaux de paroi, sporopollénine, pollenkitt, tryphine et nombre de protéines qui contrôlent la réaction d'incompatibilité. Tapetum contrôle également la fertilité ou la stérilité des microspores ou des grains de pollen.
Il existe deux types de tapetum en fonction de son comportement. Ils sont:
Tapetum sécrétoire (pariétal/glandulaire/ cellulaire) : Le tapetum conserve la position d'origine et l'intégrité cellulaire et nourrit les microspores en développement.
Tapetum invasif ( périplasmodium): Les cellules perdent leurs parois tangentielles et radiales internes et le protoplaste de toutes les cellules du tapetum fusionne pour former un périplasmodium.
Fonctions du Tapetum :
· Il fournit la nutrition aux microspores en développement.
· Il contribue à la sporopollénine à travers les corps ubich et joue ainsi un rôle important dans la formation de la paroi pollinique.
· La matière pollinique est apportée par les cellules du tapis et est ensuite transférée à la surface du pollen.
Les protéines de l'exine responsables de la « réaction de rejet » du stigmate sont présentes dans les cavités de l'exine. Ces protéines sont dérivées de cellules tapétales.
2. Cavité de l' anthère : La cavité de l'anthère est remplie de microspores au stade jeune ou de grains de pollen à maturité. La division méiotique des cellules mères des microspores donne naissance à des microspores de nature haploïde.
3. Conjonctif : C'est la colonne de tissu stérile entourée par le lobe de l'anthère. Il possède des tissus vasculaires. Il contribue également au tapetum interne.
Microspores et grains de pollen
Les microspores sont le produit immédiat de la méiose de la cellule mère des microspores tandis que le grain de pollen est dérivé de la microspore. Les microspores ont un protoplaste entouré d'une paroi qui n'est pas encore complètement développée. Le protoplaste du pollen est constitué d'un cytoplasme dense avec un noyau situé au centre. La paroi est différenciée en deux couches, à savoir, la couche interne appelée intine et la couche externe appelée exine. L'intine est fine, uniforme et se compose de pectine, d'hémicellulose, de cellulose et de callose ainsi que de protéines. L'exine est épaisse et se compose de cellulose, de sporopollénine et de pollenkitt. L'exine n'est pas uniforme et est mince à certains endroits. Lorsque ces zones minces sont petites et rondes, elles sont appelées pores germinatifs ou lorsqu'elles sont allongées, elles sont appelées sillons. Elle est associée à la germination des grains de pollen. La sporopollénine est généralement absente dans les pores du germe. La surface de l'exine est soit lisse, soit sculptée selon divers motifs (en forme de tige, rainuré, verruqueux, ponctué, etc.). Le motif de sculpture est utilisé dans l'identification et la classification des plantes.
La forme d'un grain de pollen varie d'une espèce à l'autre. Il peut être globuleux, ellipsoïde, fusiforme, lobé, angulaire ou en forme de croissant. La taille du pollen varie de 10 micromètres chez Myosotis à 200 micromètres chez les membres de la famille des Cucurbitacées et des Nyctaginacées
Le matériau de la paroi, la sporopollénine, est apporté à la fois par le cytoplasme du pollen et le tapetum. Il est dérivé des caroténoïdes. Il résiste à la décomposition physique et biologique. Il aide à supporter les températures élevées et résiste à l'action des acides forts, des alcalis et des enzymes. Par conséquent, il préserve le pollen pendant de longues périodes dans les gisements fossiles, et il protège également le pollen pendant son voyage de l'anthère au stigmate.
Pollenkitt est apporté par le tapetum et de couleur jaune ou orange et est principalement composé de caroténoïdes ou de flavonoïdes. C'est une couche huileuse formant une épaisse couche visqueuse sur la surface du pollen. Il attire les insectes et protège des dommages causés par les rayons UV.
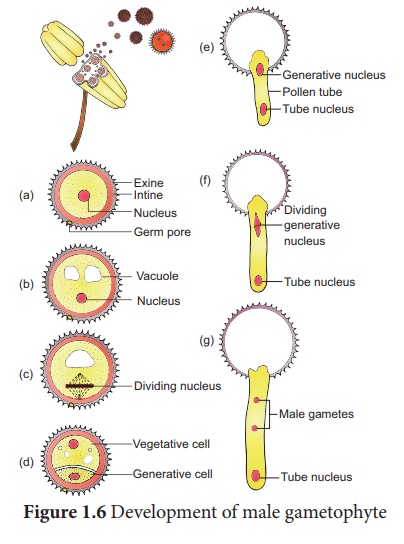
Développement du gamétophyte mâle :
La microspore est la première cellule du gamétophyte mâle et est haploïde. Le développement du gamétophyte mâle a lieu alors qu'il est encore dans le microsporange. Le noyau de la microspore se divise pour former un végétatif et un génératif noyau. Une paroi est posée autour du noyau génératif entraînant la formation de deux cellules inégales, un gros noyau irrégulier portant une réserve alimentaire abondante appelée cellule végétative et une petite cellule générative. A ce stade 2 cellules, les pollens sont libérés de l'anthère. Chez certaines plantes, la cellule générative subit à nouveau une division pour former deux gamètes mâles. Chez ces plantes, le pollen est libéré au stade 3 cellules. Dans 60% des angiospermes, le pollen est libéré au stade 2 cellules. De plus, la croissance du gamétophyte mâle ne se produit que si le pollen atteint le bon stigmate. Le pollen en atteignant le stigmate absorbe l'humidité et gonfle. L'intine se développe comme un tube pollinique à travers le pore du germe. Dans le cas où le pollen est libéré au stade 2 cellules, la cellule générative se divise dans le pollen en 2 cellules mâles (spermatozoïdes) après avoir atteint le stigmate ou dans le tube pollinique avant d'atteindre le sac embryonnaire. Les étapes du développement du gamétophyte mâle sont présentées à la figure 1.6.