La fusion des gamètes mâles et femelles est appelée fécondation . La fécondation chez les angiospermes est de type double fécondation .
Événements de fécondation
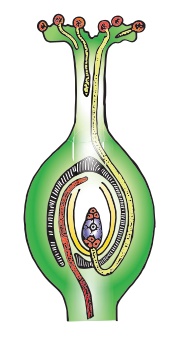
Les étapes impliquées dans la double fécondation sont :- la germination du pollen pour former le tube pollinique dans le stigmate ; croissance du tube pollinique dans le style; direction du tube pollinique vers le micropyle de l'ovule ; entrée du tube pollinique dans l'un des synergides du sac embryonnaire, écoulement des gamètes mâles; syngamie et triple fusion. Les événements depuis le dépôt de pollen sur le stigmate jusqu'à l'entrée du tube pollinique dans l'ovule sont appelés interaction pollen-pistil. C'est un processus dynamique qui consiste à reconnaître le pollen et à favoriser ou inhiber sa germination et sa croissance.
Pollen sur la stigmatisation
Dans la nature, une variété de pollens tombent sur le stigmate réceptif, mais tous ne germent pas et n'engendrent pas la fécondation. La surface réceptive du stigmate reçoit le pollen. Si le pollen est compatible avec le stigmate, il germe pour former un tube. Ceci est facilité par le fluide stigmatique dans le stigmate humide et la pellicule dans le stigmate sec . Ces deux décident également de l'incompatibilité et de la compatibilité du pollen grâce à une réaction protéique de reconnaissance-rejet entre le pollen et la surface du stigmate. Une incompatibilité sexuelle peut exister entre différentes espèces (interspécifique) ou entre membres d'une même espèce (intraspécifique). Cette dernière est appelée auto-incompatibilité. Le premier changement visible dans le pollen, peu de temps après son arrivée sur le stigmate, est l'hydratation. Les protéines de la paroi pollinique sont libérées de la surface. Au cours de la germination du pollen, tout son contenu se déplace dans le tube pollinique. La croissance est limitée à la pointe du tube et tout le contenu cytoplasmique se déplace vers la région de la pointe. La partie restante du tube pollinique est occupée par une vacuole qui est coupée de la pointe par un bouchon calleux. L'extrémité extrême du tube pollinique apparaît hémisphérique et transparente lorsqu'elle est observée au microscope. C'est ce qu'on appelle le bloc de capuchon. Dès que le bloc de capuchon disparaît, la croissance du tube pollinique s'arrête.
Tube à pollen dans le style
Après la germination, le tube pollinique entre dans le style à partir du stigmate. La croissance du tube pollinique dans le style dépend du type de style.
Types de styles
Il existe trois types de style a) style creux ou ouvert b) style plein ou style fermé c) style semi-plein ou semi-fermé.
Style creux (style ouvert) : Il est courant chez les monocotylédones. Un canal creux allant du stigmate à la base du style est présent. Le canal est tapissé d'une seule couche de cellules du canal glandulaire (tissu de transmission). Elles sécrètent des substances mucilagineuses. Le tube pollinique se développe à la surface des cellules tapissant le canal stylaire. Le canal est rempli de sécrétions qui servent de nutrition pour la croissance des tubes polliniques et contrôlent également la réaction d'incompatibilité entre le style et le tube pollinique. Les sécrétions contiennent des glucides, des lipides et certaines enzymes comme les estérases, les phosphatases acides ainsi que des protéines contrôlant la compatibilité.
Style solide (type fermé) : Il est courant chez les dicotylédones. Il se caractérise par la présence d'un noyau central de cellules allongées hautement spécialisées appelées tissus de transmission. Ceci est équivalent aux cellules de revêtement de style creux et remplit la même fonction. Son contenu est également similaire au contenu de ces cellules. Le tube pollinique se développe à travers les espaces intercellulaires du tissu de transmission.
Style semi-solide (type semi-fermé) : C'est intermédiaire entre le type solide et ouvert.
Il existe une divergence d'opinion sur la nature du tissu de transmission. Certains auteurs considèrent qu'il ne se trouve que dans les styles solides tandis que d'autres considèrent que les cellules de revêtement du style creux ont également des tissus de transmission.
Entrée du tube pollinique dans l'ovule : Il existe trois types d'entrée du tube pollinique dans l'ovule (Figure 1.18).
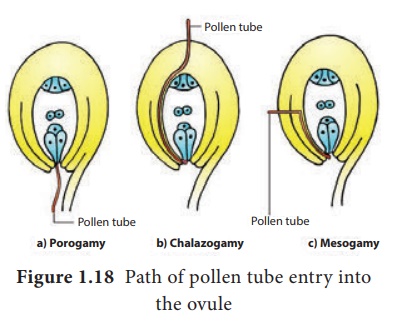
Porogamie : lorsque le tube pollinique pénètre par le micropyle.
Chalazogamie : lorsque le tube pollinique pénètre par la chalaze.
Mésogamie : lorsque le tube pollinique pénètre par le tégument.
Entrée du tube pollinique dans le sac embryonnaire :
Quel que soit le lieu d'entrée du tube pollinique dans l'ovule, il pénètre dans le sac embryonnaire à l'extrémité micropylaire. Le pollen entre dans le sac embryonnaire directement dans l'un des synergides.
La croissance du tube pollinique vers l'ovaire, l'ovule et le sac embryonnaire est due à la présence de substances chimiotropes. Le tube pollinique après avoir parcouru toute la longueur du style pénètre dans la loge de l'ovaire où il est guidé vers le micropyle de l'ovule par une structure appelée obturateur (Voir Connaissez-vous). Après avoir atteint le sac embryonnaire, un pore se forme dans la paroi du tube pollinique à son sommet ou juste derrière le sommet. Le contenu du tube pollinique (deux gamètes mâles, le noyau végétatif et le cytoplasme) est déchargé dans les synergides dans lesquels pénètre le tube pollinique. Le tube pollinique ne se développe pas au-delà, dans le sac embryonnaire. Le noyau du tube se désorganise.
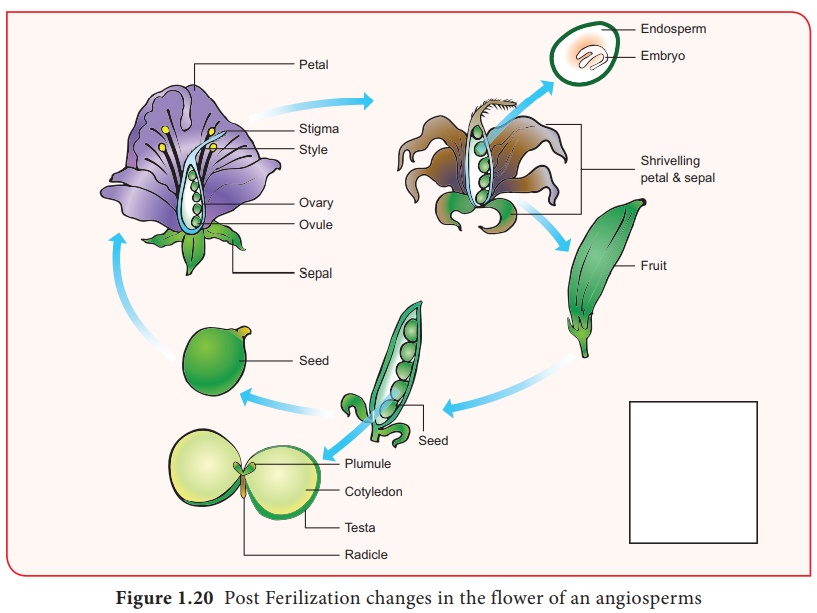
Double fécondation et triple fusion
SG Nawaschin et L.Guignard en 1898 et 1899, ont observé chez Lilium et Fritillaria que les deux gamètes mâles libérés d'un gamétophyte mâle sont impliqués dans la fécondation. Ils fertilisent deux composants différents du sac embryonnaire. Étant donné que les deux gamètes mâles sont impliqués dans la fécondation, le phénomène est appelé double fécondation et est unique aux angiospermes. L'un des gamètes mâles fusionne avec le noyau de l'œuf (syngamie) pour former le Zygote. (Figure 1.19)
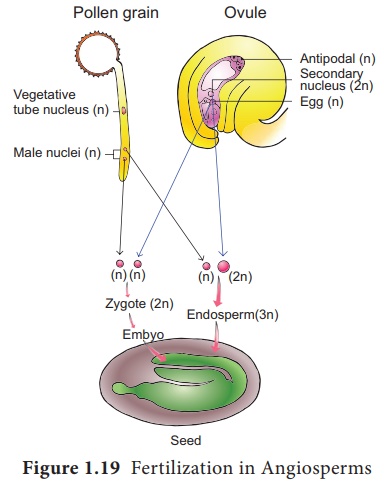
Le deuxième gamète migre vers la cellule centrale où il fusionne avec les noyaux polaires ou leur produit de fusion, le noyau secondaire et forme le noyau primaire de l'endosperme (PEN) .
Puisqu'il s'agit de la fusion de trois noyaux, ce phénomène est appelé triple fusion . Cet acte entraîne la formation de l'endosperme qui constitue le tissu nutritif de l'embryon.
Plus à savoir
· Le réceptacle devient charnu et comestible autour du fruit renfermant les graines comme dans Pyrus malus (pomme)
· Le calice peut persister et s'agrandir ( Solanum melongena) ou recouvrir le fruit (Physalis minima )
· La tige florale ou l'axe sous le gynécée s'agrandit en un corps juteux en forme de poire qui est comestible ( Anacardium occidentale).

Le Périanthe devient charnu comme dans le fruit Jack.
Les cellules présentes à l'extrémité du tégument externe autour du micropyle se développent en une structure charnue appelée caroncule . ( Ricinus communis).
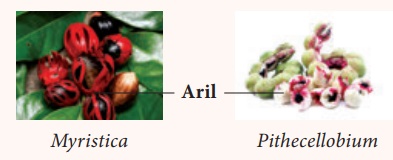
• Le funicule se développe en une structure charnue souvent très colorée et appelée arille . ( Myristica et Pithecellobium )

• Le tissu nucellaire est soit complètement absorbé par le sac embryonnaire en développement et l'embryon ou une petite partie peut rester comme tissu de stockage. Ainsi, le reste de tissu nucellaire dans la graine est appelé périsperme . Exemple : poivre noir et betterave